


COVID-19 Deep Dive Part II: Vaccine Complications
Our governments are compelling us to inject poison into our bodies

The Historical Failure of SARS Vaccines
Ever since SARS-CoV was discovered, vaccine researchers have attempted to develop a SARS vaccine. For two decades, this research has failed to produce any results.
One SARS vaccine study dated to 2012 shows the results of employing an inactivated whole virus or virus-like particle vaccine in a murine model. The animals developed Th2 immunopathology and eosinophilia in their lungs upon challenge with the live virus.
All vaccines induced serum neutralizing antibody with increasing dosages and/or alum significantly increasing responses. Significant reductions of SARS-CoV two days after challenge was seen for all vaccines and prior live SARS-CoV. All mice exhibited histopathologic changes in lungs two days after challenge including all animals vaccinated (Balb/C and C57BL/6) or given live virus, influenza vaccine, or PBS suggesting infection occurred in all. Histopathology seen in animals given one of the SARS-CoV vaccines was uniformly a Th2-type immunopathology with prominent eosinophil infiltration, confirmed with special eosinophil stains. The pathologic changes seen in all control groups lacked the eosinophil prominence.
Eosinophils are a part of a subcategory of white blood cells known as granulocytes. The three main types of granulocytes found in the blood are neutrophils, eosinophils, and basophils, with mast cells additionally found in the tissues.
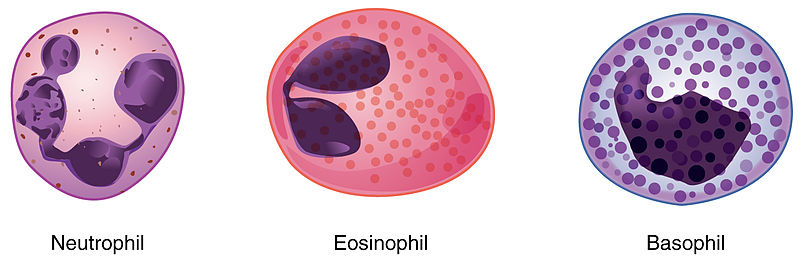
Granulocytes fight infection by deploying granules containing various kinds of destructive enzymes. As described in Part I of this series, neutrophils aggressively fight pathogens by dispersing superoxide dismutase and myeloperoxidase to make peroxide and bleach, denaturing and destroying pathogens. They also engulf pathogens and expose them to these enzymes, sadistically dissolving them. It’s actually surprisingly violent. Neutrophil extracellular trap formation has no macro-scale analogy that doesn’t sound ridiculous. Neutrophils actually eject destructive enzymes along with histones and their own nuclear DNA when they do it. Picture a man drinking acid, and then screaming and ripping his acidic guts out and throwing them at you to kill you.
Eosinophils are specialized in mediating allergic reactions and in triggering immune reactions to parasitic infections. They patrol around and release inflammatory cytokines when they spot something amiss.
Basophils are the rarest type, their function incompletely understood by science. It is known that they contain heparin, histamines, and serotonin, and may have an immune mediating purpose.
Mature granulocytes are terminally differentiated and do not divide; their immature progenitors divide and replicate in bone marrow.
Mature granulocytes are a little bit something like the Sardaukar:
They are fanatical, face-painted warriors who are well prepared for the inevitability of death.
The word “inflammation” basically means something that summons these cells (along with the various cells of the adaptive immune system). When inflammation is systemic and leads to organ failure, we refer to it as sepsis, or systemic inflammatory response syndrome (SIRS). Sepsis is no joke. It puts people in the ICU all the time and is very difficult to treat.
Yes, your own immune system is armed to the teeth and perfectly capable of killing you.
Innate and Adaptive Immune Systems
The immune system is split into two general parts. The innate immune system, and the adaptive immune system. All responses to infection with a pathogen proceed in the same general manner; first, damaged cells give off danger signals and produce inflammatory cytokines. These cytokines act as a smoke signal to draw in the aforementioned face-painted fanatical warriors, the white blood cells. Neutrophils attack the pathogen as best they can with destructive enzymes, macrophages follow after them to engulf and dissolve the debris, and your own cells cower beside this awesome display of firepower, desperately trying to survive by using antioxidant enzymes and survival pathways to detoxify themselves of the antiseptic soup of granules the immune cells are putting out.
However, the purpose of this initial counter-assault is only to stall the pathogen while the studious strategists of the adaptive immune system, the T and B lymphocytes and dendritic cells, come up with a more permanent solution in the form of antibodies. Once the body successfully begins making antibodies, those antibodies latch onto the pathogen’s external proteins and their binding sites like little limpet mines and macrophages make a beeline for the neutralized pathogens to engulf and destroy them, shutting down the infection for good.
The purpose of a vaccine, generally speaking, is to induce an adaptive immune response without the drawbacks of a full-blown infection. The way this is done is by using whole killed or live attenuated virus, or protein subunits from that virus, to present antigens to the immune system and stimulate an antibody response. Vaccines serve no other purpose whatsoever. The only point of a vaccine is to get an adaptive immune response without an infection or inflammation, by giving the immune system a training target so it recognizes the real pathogen on sight.
If a vaccine produces antibodies that do not grant immunity to a pathogen, it’s useless. If a vaccine fails to generate immunity and actually stimulates an inflammatory response that damages tissue, it’s worse than useless.
Types of Vaccines
There are four main types of vaccines; Live Attenuated Vaccine (LAV), Inactivated, Toxoid, and Subunit.
Live attenuated vaccines use a weakened virus, inactivated vaccines use a killed virus, toxoid vaccines use an inactivated toxin from a pathogen, and subunit vaccines are protein-based. Very often, these vaccines include various toxic adjuvants that stimulate a stronger immune response to the antigens presented in the vaccine.
A virus is, fundamentally, a collection of proteins embedded in a membrane (or not; non-enveloped “naked” viruses without a lipid membrane do exist), encasing a genome and acting as a delivery mechanism for that genome, introducing it into live cells to produce more virions. The main target antigens of a virus are usually the outer structural proteins it uses to bind to and fuse with living cells. In SARS-CoV-2, this would be the Spike, for instance.
In recent years, two new types of vaccines have appeared: viral vector vaccines and mRNA vaccines. These vaccines use a completely different approach; the human-cell-as-bioreactor concept. Basically, they deliver genetic material to human cells to get them to manufacture viral proteins and present them on their surfaces, which, in turn, is supposed to stimulate an immune response. The first viral vector vaccines for Ebola were made available in the 2010s, using Vesicular Stomatitis Virus as the vector.
For COVID-19, a number of adenovirus vector-based vaccines were developed, such as the Janssen/J&J COVID-19 Vaccine Ad.26.COV2.S, the AstraZeneca COVID-19 Vaccine known by the trade names Vaxzevria or Covishield, and Gamaleya’s Sputnik-V. These are joined by two mRNA vaccines: Moderna’s Spikevax/mRNA-1273, and Pfizer-BioNTech’s Comirnaty/BNT162B2.
Technically, it is not necessary to do this using living cells in the body. It is entirely possible to produce a subunit vaccine for SARS-CoV-2 by inserting the gene for SARS-CoV-2 Spike into, for instance, E. Coli or Spirulina or some other bacteria, culturing it in a bioreactor vat, and then collecting and purifying the protein. This is how recombinant insulin and various other biologics are made.
With viral vector and mRNA vaccines, the vaccine is genetic code, albeit delivered by two different mechanisms. The muscle cells in the recipient’s deltoid are the bioreactors. Viral vector vaccines introduce genetic material into the cells by way of a replication-defective recombinant adenovirus, and mRNA vaccines introduce messenger RNA into the cells using synthetic lipid nanoparticles filled with messenger RNA that fuse with the cells and release their cargo.
The COVID-19 pandemic marked the first-ever use of viral vector and mRNA vaccines on a very large scale. These vaccines were rushed through highly accelerated trials and sold to governments under purchase agreements that shielded their manufacturers from any liability for their failure.
Making an Inert Spike and Bypassing the Immune System
SARS-CoV-2 Spike is a very nasty viral glycoprotein. Its entire purpose is to fuse with human ACE2, shed its trimeric heads, and then pull the virus and the cell’s membranes together. It does so using molecular mechanics on a tiny scale, like the hydraulic arm of an excavator. Before it could be used as a vaccine antigen, it had to be made inert. The way researchers proposed to do this was by making proline substitutions in the Spike’s sequence, locking the trimeric heads in the prefusion conformation. This research resulted in prefusion-locked variants of the Spike such as 2P and HexaPro:
bioRxiv (Preprint) - Structure-based Design of Prefusion-stabilized SARS-CoV-2 Spikes
The COVID-19 pandemic caused by the novel coronavirus SARS-CoV-2 has led to accelerated efforts to develop therapeutics, diagnostics, and vaccines to mitigate this public health emergency. A key target of these efforts is the spike (S) protein, a large trimeric class I fusion protein that is metastable and difficult to produce recombinantly in large quantities. Here, we designed and expressed over 100 structure-guided spike variants based upon a previously determined cryo-EM structure of the prefusion SARS-CoV-2 spike. Biochemical, biophysical and structural characterization of these variants identified numerous individual substitutions that increased protein yields and stability. The best variant, HexaPro, has six beneficial proline substitutions leading to ~10-fold higher expression than its parental construct and is able to withstand heat stress, storage at room temperature, and multiple freeze-thaws. A 3.2 Å-resolution cryo-EM structure of HexaPro confirmed that it retains the prefusion spike conformation. High-yield production of a stabilized prefusion spike protein will accelerate the development of vaccines and serological diagnostics for SARS-CoV-2.
The assumption, generally speaking, is that locking the Spike in the prefusion conformation allows it to be presented to the immune system as a viable vaccine antigen, but prevents it from having any toxic or cytopathic effects by unwanted activation and fusion.
The Spike also has a transmembrane domain in its tail, in the S2 subunit, so it would become a membrane-bound protein once expressed by deltoid muscle cells. This would keep it from being exported by those cells and migrating all over the body, theoretically.
With a size of 180–200 kDa, the S protein consists of an extracellular N-terminus, a transmembrane (TM) domain anchored in the viral membrane, and a short intracellular C-terminal segment [11]. S normally exists in a metastable, prefusion conformation; once the virus interacts with the host cell, extensive structural rearrangement of the S protein occurs, allowing the virus to fuse with the host cell membrane. The spikes are coated with polysaccharide molecules to camouflage them, evading surveillance of the host immune system during entry [12].
This Spike was then encoded into DNA for the viral vector vaccines and mRNA for the lipid nanoparticle vaccines. For the mRNA vaccines, some means of stabilizing the mRNA and evading immune detection was necessary. DNA and Messenger RNA are made up of long chains of five different nucleobases; adenine (A), cytosine (C), guanine (G), thymine (T), and uracil (U). For the mRNA vaccines, they took uracil and replaced it with pseudouridylyl (Ψ). This enabled it to evade detection by toll-like receptors, a type of pattern recognition receptor which triggers inflammation upon activation.
Pseudouridylation has another effect; it makes mRNA molecularly stable and causes it to resist degradation.
Frontiers - The Critical Contribution of Pseudouridine to mRNA COVID-19 Vaccines
While the changes seem subtle (in fact, Ψ can base-pair with adenosine just as uridine does), Ψ can alter RNA structure in a relatively significant way, mainly by improving base-pairing, base stacking, and contributing to making the backbone more rigid (through a network of hydrogen bonding interactions) (Davis, 1995; Charette and Gray, 2000; Newby and Greenbaum, 2001, 2002a, 2002b). As such, RNA pseudouridylation generally stabilizes the RNA. Thus, it is not surprising that the presence of this RNA modification confers distinct biophysical and biochemical properties to the RNA. For example, Ψ favors a C3′-endo conformation in the RNA (Kierzek et al., 2014; Westhof, 2019). Further, it seems that Ψ increases the protection of the RNA against nucleases. A study from Naylor et al. showed that Ψ-containing dinucleotides were more resistant to degradation from snake venom and spleen phosphodiesterases, than the U-containing counterparts (Naylor et al., 1965).
This leads to the mRNA persisting in the body for a long period of time.
Endogenous Reverse Transcription
A common theme among fact-checkers is that the vaccines do not mess with the recipients’ DNA. The genetic material in the vaccines merely produces a protein, and then harmlessly degrades, they insist. Therefore, according to them, these vaccines do not constitute gene therapy.
Reuters - Fact Check-mRNA vaccines are distinct from gene therapy, which alters recipient’s genes
Pfizer/BioNTech and Moderna have both developed shots that use a piece of genetic code from SARS-CoV-2, the coronavirus that causes COVID-19, to prompt an immune response in recipients (here). However, experts told Reuters that this is not the same as gene therapy.
“As mRNA is genetic material, mRNA vaccines can be looked at as a genetic-based therapy, but they are classified as vaccines and are not designed to alter your genes,” said Dr Adam Taylor, a virologist and research fellow at the Menzies Health Institute, Queensland, Griffith University.
Sounds great, right? Except it’s wrong.
Here’s Stephan Oelrich stating that these vaccines are, essentially, a form of gene therapy:
And here’s a study showing that the genes delivered by the vaccines can be permanently integrated into the recipient’s genome in as little as six hours:
Preclinical studies of COVID-19 mRNA vaccine BNT162b2, developed by Pfizer and BioNTech, showed reversible hepatic effects in animals that received the BNT162b2 injection. Furthermore, a recent study showed that SARS-CoV-2 RNA can be reverse-transcribed and integrated into the genome of human cells. In this study, we investigated the effect of BNT162b2 on the human liver cell line Huh7 in vitro. Huh7 cells were exposed to BNT162b2, and quantitative PCR was performed on RNA extracted from the cells. We detected high levels of BNT162b2 in Huh7 cells and changes in gene expression of long interspersed nuclear element-1 (LINE-1), which is an endogenous reverse transcriptase. Immunohistochemistry using antibody binding to LINE-1 open reading frame-1 RNA-binding protein (ORFp1) on Huh7 cells treated with BNT162b2 indicated increased nucleus distribution of LINE-1. PCR on genomic DNA of Huh7 cells exposed to BNT162b2 amplified the DNA sequence unique to BNT162b2. Our results indicate a fast up-take of BNT162b2 into human liver cell line Huh7, leading to changes in LINE-1 expression and distribution. We also show that BNT162b2 mRNA is reverse transcribed intracellularly into DNA in as fast as 6 h upon BNT162b2 exposure.
Again, as I spelled out in previous articles, protein synthesis, one of the most basic processes of life, is as follows: DNA makes RNA makes Proteins. It really is that simple. DNA strands are permanent, long-term storage blueprints, which are copied to RNA, the working form of a gene, and the RNA strands leave the nucleus and are translated to proteins by ribosomes that read them off and build the protein one amino acid at a time using the codons in the RNA strand as a template. Cells decide how much of a given protein to make by increasing or decreasing gene expression.
Retroviruses like HIV use reverse transcriptase to turn their genomes into DNA and integrate that DNA into the genome of the host’s cells, causing them to permanently produce HIV proteins and virions. That’s why HIV never goes away. The code to make more HIV virions becomes a permanent part of the host’s cells.
So, SARS-CoV-2 doesn’t have a reverse transcriptase and isn’t a retrovirus, and the vaccine doesn’t contain a reverse transcriptase. So what gives? Why would it integrate into the genome?
Well, as it turns out, human cells actually have their own endogenous reverse transcriptase, possessed by LINE-1 Retrotransposons:
In higher eukaryotic genomes, Long Interspersed Nuclear Element 1 (LINE-1) retrotransposons represent a large family of repeated genomic elements. They transpose using a reverse transcriptase (RT), which they encode as part of the ORF2p product. RT inhibition in cancer cells, either via RNA interference-dependent silencing of active LINE-1 elements, or using RT inhibitory drugs, reduces cancer cell proliferation, promotes their differentiation and antagonizes tumor progression in animal models. Indeed, the non-nucleoside RT inhibitor efavirenz has recently been tested in a phase II clinical trial with metastatic prostate cancer patients. An in-depth analysis of ORF2p in a mouse model of breast cancer showed ORF2p to be precociously expressed in precancerous lesions and highly abundant in advanced cancer stages, while being barely detectable in normal breast tissue, providing a rationale for the finding that RT-expressing tumors are therapeutically sensitive to RT inhibitors. We summarize mechanistic and gene profiling studies indicating that abundant LINE-1-derived RT can “sequester” RNA substrates for reverse transcription in tumor cells, entailing the formation of RNA:DNA hybrid molecules and impairing the overall production of regulatory miRNAs, with a global impact on the cell transcriptome. Based on these data, LINE-1-ORF2 encoded RT has a tumor-promoting potential that is exerted at an epigenetic level. We propose a model whereby LINE1-RT drives a previously unrecognized global regulatory process, the deregulation of which drives cell transformation and tumorigenesis with possible implications for cancer cell heterogeneity.
Oops. Now the gene from the vaccine is a part of the recipient’s genome. Which means it is gene therapy. If the region of the genome it is incorporated into is not silent, and actually expresses a protein, that cell will now start producing that protein indefinitely.
Except there’s a bit of a problem here. That gene encodes SARS-CoV-2 Spike. A highly toxic protein.
SARS-CoV-2 Spike Inoculation as a Toxic Proteinopathy
SARS-CoV-2 Spike has many motifs on its surface which are of a highly questionable nature.
J Biol Regul Homeost Agents - Be aware of SARS-CoV-2 spike protein: There is more than meets the eye
Human The COVID-19 pandemic necessitated the rapid production of vaccines aimed at the production of neutralizing antibodies against the COVID-19 spike protein required for the corona virus binding to target cells. The best well-known vaccines have utilized either mRNA or an adenovirus vector to direct human cells to produce the spike protein against which the body produces mostly neutralizing antibodies. However, recent reports have raised some skepticism as to the biologic actions of the spike protein and the types of antibodies produced. One paper reported that certain antibodies in the blood of infected patients appear to change the shape of the spike protein so as to make it more likely to bind to cells, while other papers showed that the spike protein by itself (without being part of the corona virus) can damage endothelial cells and disrupt the blood-brain barrier. These findings may be even more relevant to the pathogenesis of long-COVID syndrome that may affect as many as 50% of those infected with SARS-CoV-2. In COVID-19, a response to oxidative stress is required by increasing anti-oxidant enzymes. In this regard, it is known that polyphenols are natural anti-oxidants with multiple health effects. Hence, there are even more reasons to intervene with the use of anti-oxidant compounds, such as luteolin, in addition to available vaccines and anti-inflammatory drugs to prevent the harmful actions of the spike protein.
Although ACE2 (angiotensin converting enzyme 2) is considered the primary receptor for CoV-2 cell entry, recent reports suggest that alternative pathways may contribute. This paper considers the hypothesis that viral binding to cell-surface integrins may contribute to the high infectivity and widespread extra-pulmonary impacts of the SARS-CoV-2 virus. This potential is suggested on the basis of the emergence of an RGD (arginine-glycine-aspartate) sequence in the receptor-binding domain of the spike protein. RGD is a motif commonly used by viruses to bind cell-surface integrins. Numerous signaling pathways are mediated by integrins and virion binding could lead to dysregulation of these pathways, with consequent tissue damage. Integrins on the surfaces of pneumocytes, endothelial cells and platelets may be vulnerable to CoV-2 virion binding. For instance, binding of intact virions to integrins on alveolar cells could enhance viral entry. Binding of virions to integrins on endothelial cells could activate angiogenic cell signaling pathways; dysregulate integrin-mediated signaling pathways controlling developmental processes; and precipitate endothelial activation to initiate blood clotting. Such a procoagulant state, perhaps together with enhancement of platelet aggregation through virions binding to integrins on platelets, could amplify the production of microthrombi that pose the threat of pulmonary thrombosis and embolism, strokes and other thrombotic consequences. The susceptibility of different tissues to virion–integrin interactions may be modulated by a host of factors, including the conformation of relevant integrins and the impact of the tissue microenvironment on spike protein conformation. Patient-specific differences in these factors may contribute to the high variability of clinical presentation. There is danger that the emergence of receptor-binding domain mutations that increase infectivity may also enhance access of the RGD motif for integrin binding, resulting in viral strains with ACE2 independent routes of cell entry and novel integrin-mediated biological and clinical impacts. The highly infectious variant, B.1.1.7 (or VUI 202012/01), includes a receptor-binding domain amino acid replacement, N501Y, that could potentially provide the RGD motif with enhanced access to cell-surface integrins, with consequent clinical impacts.
The post-infection of COVID-19 includes a myriad of neurologic symptoms including neurodegeneration. Protein aggregation in brain can be considered as one of the important reasons behind the neurodegeneration. SARS-CoV-2 Spike S1 protein receptor binding domain (SARS-CoV-2 S1 RBD) binds to heparin and heparin binding proteins. Moreover, heparin binding accelerates the aggregation of the pathological amyloid proteins present in the brain. In this paper, we have shown that the SARS-CoV-2 S1 RBD binds to a number of aggregation-prone, heparin binding proteins including Aβ, α-synuclein, tau, prion, and TDP-43 RRM. These interactions suggests that the heparin-binding site on the S1 protein might assist the binding of amyloid proteins to the viral surface and thus could initiate aggregation of these proteins and finally leads to neurodegeneration in brain. The results will help us to prevent future outcomes of neurodegeneration by targeting this binding and aggregation process.
SARS-CoV-2 may cause acute respiratory disease, but the infection can also initiate neurological symptoms. Here we show that SARS-CoV-2 infection causes brain inflammation in the macaque model. An increased metabolic activity in the pituitary gland of two macaques was observed by longitudinal positron emission tomography-computed tomography (PET-CT). Post-mortem analysis demonstrated infiltration of T-cells and activated microglia in the brain, and viral RNA was detected in brain tissues from one animal. We observed Lewy bodies in brains of all rhesus macaques. These data emphasize the virus’ capability to induce neuropathology in this nonhuman primate model for SARS-CoV-2 infection. As in humans, Lewy body formation is an indication for the development of Parkinson’s disease, this data represents a warning for potential long-term neurological effects after SARS-CoV-2 infection.
Nature Neuroscience - The S1 protein of SARS-CoV-2 crosses the blood–brain barrier in mice
It is unclear whether severe acute respiratory syndrome coronavirus 2, which causes coronavirus disease 2019, can enter the brain. Severe acute respiratory syndrome coronavirus 2 binds to cells via the S1 subunit of its spike protein. We show that intravenously injected radioiodinated S1 (I-S1) readily crossed the blood–brain barrier in male mice, was taken up by brain regions and entered the parenchymal brain space. I-S1 was also taken up by the lung, spleen, kidney and liver. Intranasally administered I-S1 also entered the brain, although at levels roughly ten times lower than after intravenous administration. APOE genotype and sex did not affect whole-brain I-S1 uptake but had variable effects on uptake by the olfactory bulb, liver, spleen and kidney. I-S1 uptake in the hippocampus and olfactory bulb was reduced by lipopolysaccharide-induced inflammation. Mechanistic studies indicated that I-S1 crosses the blood–brain barrier by adsorptive transcytosis and that murine angiotensin-converting enzyme 2 is involved in brain and lung uptake, but not in kidney, liver or spleen uptake.
SARS-CoV-2 Spike’s S1 subunit can cross the blood-brain barrier and induce Lewy body formation. If it can do this to someone who’s infected, then it’s an open question whether or not a vaccine that produces this protein in the body can do the same. It doesn’t just have an ACE2-binding region; it also has integrin and heparin-binding regions that may be exposed even in prefusion-locked Spike.
As a matter of fact, SARS-CoV-2 Spike permeabilizes the BBB:
Springer - SARS-CoV-2 Spike Protein Disrupts Blood–Brain Barrier Integrity via RhoA Activation
ELISA assays indicated that S1 spike protein significantly increased the activation of RhoA, demonstrating that the small GTPase influences barrier breakdown in response to SARS-CoV-2. The activation of RhoA has been shown to induce cell contractility and cytoskeleton restructuring, resulting in enhanced cell motility and disrupted barrier integrity (Shaw et al. 1998; Mikelis et al. 2015). This conclusion is supported by permeability and TEER testing showing that the detrimental effects of S1 spike protein on the BBB are ablated by inhibiting RhoA activation (Fig. 3). Given that RhoA activates the Rho kinase (ROCK), it is noteworthy that the therapeutic effects of ROCK inhibition have already been considered for the treatment of severe COVID-19 (Abedi et al. 2020b). In fact, preclinical studies have shown the benefits of inhibiting the Rho-ROCK pathway to improve pulmonary outcomes (Xu et al. 2019; Abedi et al. 2020a). Additionally, ROCK inhibitors have been shown to protect pulmonary tissue during severe respiratory illness (Abedi et al. 2020a, b). The effects of ROCK inhibition seems to also confer vascular protection by showing improved neurological outcomes following ischemic stroke (Shibuya et al. 2005). Perhaps a similar therapeutic approach could be developed to prevent neurological deficits associated with COVID-19.
But wait, you say! Isn’t the protein locked to the cell surface by the transmembrane domain? Yes, in theory. Except the SARS-CoV-2 Spike is made up of different parts; the S1 and S2 subunits. Human proteases, such as TMPRSS2, may cleave S1 from S2, causing S1 to float away into the extracellular space and perhaps even enter the bloodstream. From there, it may cross the blood-brain barrier, and then cause amyloidogenic effects in the brain. This is just one possible mechanism of injury. There are others.
SARS-CoV-2 Spike has a superantigenic region, also known as a SAg.
The binding epitope on S harbors a sequence motif unique to SARS-CoV-2 (not present in other SARS-related coronaviruses), which is highly similar in both sequence and structure to the bacterial superantigen staphylococcal enterotoxin B. This interaction between the virus and human T cells could be strengthened by a rare mutation (D839Y/N/E) from a European strain of SARS-CoV-2. Furthermore, the interfacial region includes selected residues from an intercellular adhesion molecule (ICAM)-like motif shared between the SARS viruses from the 2003 and 2019 pandemics. A neurotoxin-like sequence motif on the receptor-binding domain also exhibits a high tendency to bind TCRs. Analysis of the TCR repertoire in adult COVID-19 patients demonstrates that those with severe hyperinflammatory disease exhibit TCR skewing consistent with superantigen activation. These data suggest that SARS-CoV-2 S may act as a superantigen to trigger the development of MIS-C as well as cytokine storm in adult COVID-19 patients, with important implications for the development of therapeutic approaches.
Anti-Spike antibodies can behave like autoantibodies and attack healthy tissue:
Taken together, the results of the in vitro assay indicated that certain antibodies specific to the spike proteins of the COVID-19 and SARS-CoV viruses have the potential to mislead the immune system to attack the host by binding to sick cells such as human lung epithelium cells in vivo. We termed this mechanism of action of the antibodies as “Antibody Dependent Auto-Attack (ADAA)”.
The Spike itself is capable of triggering all kinds of potentially harmful intracellular activity by acting as a ligand for various types of receptors:
The world is suffering from the coronavirus disease 2019 (COVID-19) pandemic caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). SARS-CoV-2 uses its spike protein to enter the host cells. Vaccines that introduce the spike protein into our body to elicit virus-neutralizing antibodies are currently being developed. In this article, we note that human host cells sensitively respond to the spike protein to elicit cell signaling. Thus, it is important to be aware that the spike protein produced by the new COVID-19 vaccines may also affect the host cells. We should monitor the long-term consequences of these vaccines carefully, especially when they are administered to otherwise healthy individuals. Further investigations on the effects of the SARS-CoV-2 spike protein on human cells and appropriate experimental animal models are warranted.
Lastly, and most appallingly of all, SARS-CoV-2 Spike has been found to localize in cell nuclei and inhibit V(D)J recombination:
MDPI - SARS–CoV–2 Spike Impairs DNA Damage Repair and Inhibits V(D)J Recombination In Vitro
Severe acute respiratory syndrome coronavirus 2 (SARS–CoV–2) has led to the coronavirus disease 2019 (COVID–19) pandemic, severely affecting public health and the global economy. Adaptive immunity plays a crucial role in fighting against SARS–CoV–2 infection and directly influences the clinical outcomes of patients. Clinical studies have indicated that patients with severe COVID–19 exhibit delayed and weak adaptive immune responses; however, the mechanism by which SARS–CoV–2 impedes adaptive immunity remains unclear. Here, by using an in vitro cell line, we report that the SARS–CoV–2 spike protein significantly inhibits DNA damage repair, which is required for effective V(D)J recombination in adaptive immunity. Mechanistically, we found that the spike protein localizes in the nucleus and inhibits DNA damage repair by impeding key DNA repair protein BRCA1 and 53BP1 recruitment to the damage site. Our findings reveal a potential molecular mechanism by which the spike protein might impede adaptive immunity and underscore the potential side effects of full-length spike-based vaccines.
V(D)J recombination is not only one of many processes vital for DNA repair, it is also essential for T and B lymphocyte formation, where recombination is used to create lymphocytes specialized for specific pathogens.
PLOS Biology - V(D)J Recombination and the Evolution of the Adaptive Immune System
The immune system needs to be able to identify and ultimately destroy foreign invaders. To do so, it utilizes two major types of immune cells, T cells and B cells (or, collectively, lymphocytes). Lymphocytes display a large variety of cell surface receptors that can recognize and respond to an unlimited number of pathogens, a feature that is the hallmark of the “adaptive” immune system. To react to such a variety of invaders, the immune system needs to generate vast numbers of receptors. If the number of different types of receptors present on lymphocytes were encoded by individual genes, the entire human genome would have to be devoted to lymphocyte receptors. To establish the necessary level of diversity, B- and T-cell receptor (BCR and TCR, respectively) genes are created by recombining preexisting gene segments. Thus, different combinations of a finite set of gene segments give rise to receptors that can recognize unlimited numbers of foreign invaders. This is accomplished by a supremely well-coordinated set of reactions, starting with cleaving DNA within specific, well-conserved recombination signal sequences (RSSs). This highly regulated step is carried out by the lymphocyte-specific recombinationactivating genes (RAG1 and RAG2). The segments are then reassembled using a common cellular repair mechanism.
Inhibiting BRCA1 and 53BP1 recruitment would have a similar effect, overall, to what RAG deficiency does in Omenn Syndrome. It would lead to lymphopenia and immunodeficiency.
Remember what that one humble chiropractor Nathan Thompson said last year about vaccinated people’s blood tests?
DR. NATHAN THOMPSON TESTS PATIENT'S IMMUNE SYSTEM AFTER EACH JAB
Decimated lymphocytes, including natural killers (NKs), would not only make one more susceptible to all viral, bacterial, fungal, or parasitic infections, it would also increase one’s chances of getting cancer.
Fact-checkers at Reuters responding to multiple social media postings about “VAIDS” have stated that there is no possible way for a vaccine to cause immunodeficiency.
Claims that there are cases of AIDS caused by vaccination, or “VAIDS” as online users claim, however, are unfounded.
“There is no phenomenon that I know of ‘Vaccine-induced immunodeficiency syndrome.’ It is not a real syndrome,” Donna Farber, chief of the Division of Surgical Sciences and Professor of Microbiology & Immunology at Columbia University, told Reuters via email.
Likewise, Stephen Gluckman, MD, a professor of Infectious Diseases in the Perelman School of Medicine at the University of Pennsylvania and the medical director of Penn Global Medicine, told Reuters “VAIDS” is “absolutely not” a real condition.
And yet, there are files that describe how Spike may prevent T and B lymphocyte formation by cutting them off right at the source.
This is the exact opposite of what you want in a vaccine. The point of a vaccine, again, is to train the adaptive immune system, not sabotage it.
Toxicity From Vaccine Components
As it turns out, pseudouridylation of vaccine mRNA might not just evade toll-like receptors (TLRs), it may actually suppress all of their activity. This is bad because TLRs are necessary to detect danger signals associated with infection and cancer.
The mRNA-based BNT162b2 vaccine from Pfizer/BioNTech was the first registered COVID-19 vaccine and has been shown to be up to 95% effective in preventing SARS-CoV-2 infections. Little is known about the broad effects of the new class of mRNA vaccines, especially whether they have combined effects on innate and adaptive immune responses. Here we confirmed that BNT162b2 vaccination of healthy individuals induced effective humoral and cellular immunity against several SARS-CoV-2 variants. Interestingly, however, the BNT162b2 vaccine also modulated the production of inflammatory cytokines by innate immune cells upon stimulation with both specific (SARS-CoV-2) and non-specific (viral, fungal and bacterial) stimuli. The response of innate immune cells to TLR4 and TLR7/8 ligands was lower after BNT162b2 vaccination, while fungi-induced cytokine responses were stronger. In conclusion, the mRNA BNT162b2 vaccine induces complex functional reprogramming of innate immune responses, which should be considered in the development and use of this new class of vaccines.
The lipid nanoparticles used in the Pfizer and Moderna vaccines may trigger rare allergic reactions:
Molecular Therapy - Allergic Reactions and Anaphylaxis to LNP-Based COVID-19 Vaccines
Mortality from coronavirus disease 2019 (COVID-19) in populations at high risk, such as the elderly, certain ethnic groups (e.g., Black/African American, and Hispanic/Latino persons), obese individuals, and those with endothelial dysfunction is substantial. While treatment options are limited, vaccination against SARS-CoV-2, the virus that causes COVID-19, is the most important global strategy in controlling the pandemic. Thus, two lipid nanoparticle (LNP)-based mRNA vaccines (Pfizer-BioNTech and Moderna) against SARS-CoV-2 have already received emergency use authorization by the US Food and Drug Administration (FDA). During December 14–23 2020, after administration of 1,893,360 first doses of the Pfizer-BioNTech COVID-19 vaccine, 175 cases of severe allergic reaction were submitted to the Vaccine Adverse Event Reporting System (VAERS).
It Doesn’t Stay in the Deltoid Muscle
These vaccines spread all over the body and bioaccumulate in the organs. This was proven by a Japanese biodistribution study of the Pfizer vaccine. The Pfizer report can be viewed in 404-proof, censorship-proof form in Brave or via IPFS gateway using the following CID:
ipfs://Qmd7DB3AsyVkwzzDkWz9xYQEG3zFLXfpGRdXYnRkCbt26j
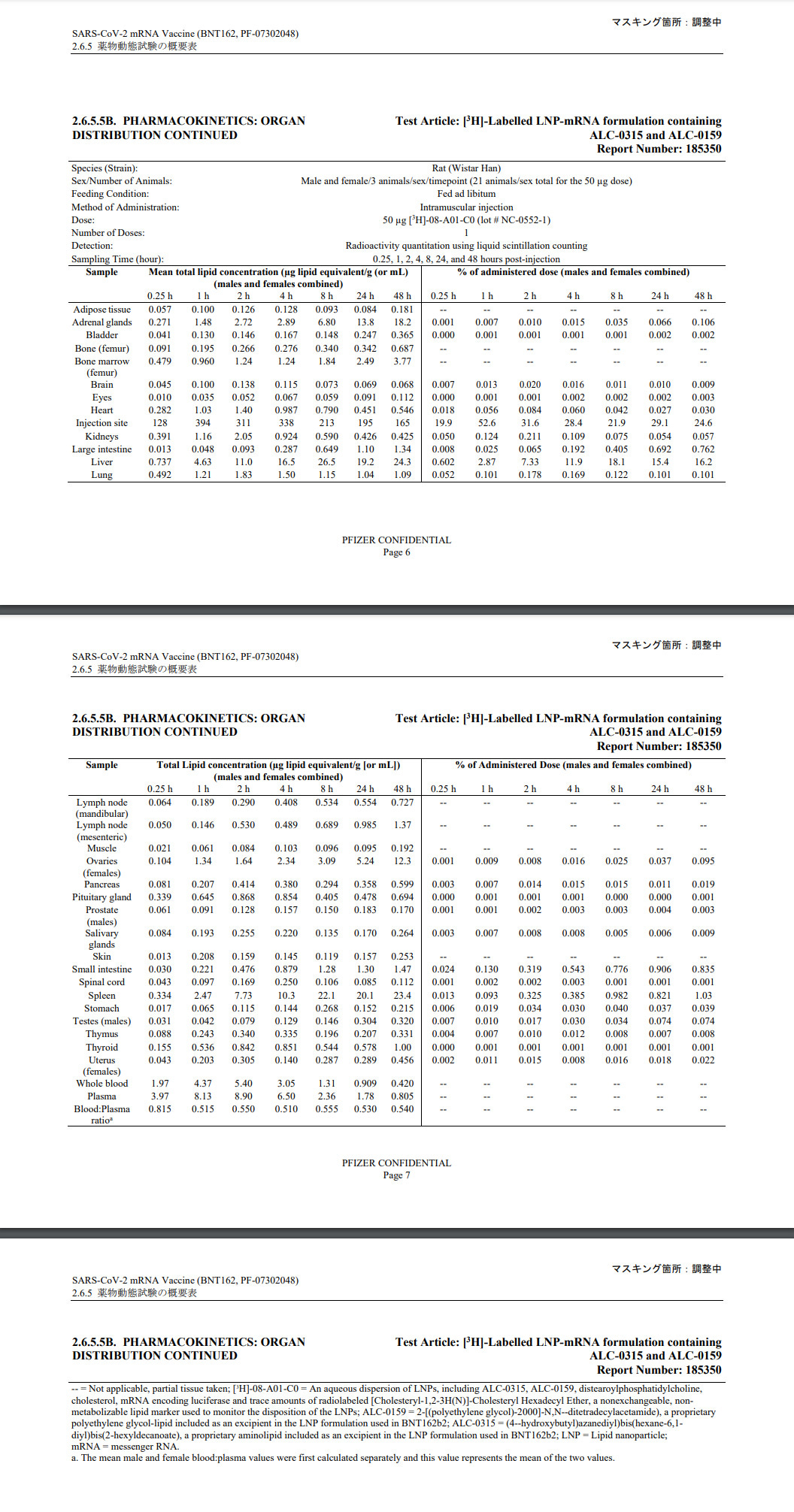
From this study, it was shown that the Pfizer-BioNTech BNT162B2 vaccine not only did not remain in the shoulder, it bioaccumulated all over the place, particularly in the gonads.
Inadvertent intravenous injection of a COVID-19 mRNA vaccine due to failure to aspirate the needle can have severe consequences:
Groups of female Balb/c mice at age of 6–8 week were given BNT162b2 COVID-19 mRNA vaccine either IV or IM, or the same volume of NS (). None of the animals showed clinical signs of lethargy, ruffled furs, hunched back posture, and rapid breathing throughout the course of observation. Significant decrease in body weight was observed in IM mRNA vaccine group (mean 3.6% ± 2.1%) starting from 1 dpi; animals recovered their initial weight at 7 dpi (). Autopsy at 1–2 dpi showed white patches over the visceral pericardium in 37.5% (1 dpi, n = 8) to 38.5% (2 dpi, n = 13) of the IV vaccine group but none in the IM vaccine or NS control groups ( and ; ; P < .05). No grossly visible changes were observed in other organs of the animals (Supplementary Figure 1).
The graphic images of dissected mice in that article showing white patches on the heart are illustrative. What’s even more tragic is that this is something that could have been prevented by the simple step of drawing back the plunger on the syringe after insertion to make sure the technician did not hit a blood vessel.
COVID-19 Vaccine-induced myocarditis is a result of a toxic vaccine combined with poor training bordering on medical negligence.
Myocarditis is not mild. Heart tissue does not readily regenerate once it has turned into scar tissue. Life expectancy after a severe case of myocarditis is measured in single-digit years. Many cases require heart transplantation to resolve.
No, it’s not your imagination. Yes, soccer players are collapsing and dying of heart failure.
Pfizer Forced to Cough Up
Recently, Pfizer were required by court order to release several highly damning documents, including this one:
https://www.scribd.com/document/544743767/5-3-6-Postmarketing-Experience?secret_password=Wvo0EDpg1BryYCC6QlWs#download
This document can also be viewed in 404-proof, censorship-proof form, using this IPFS CID:
ipfs://QmPDNfStzBca3itUn2HX4BeqQPYnivSJ672djz3eLr3gG2
This document reported tens of thousands of adverse events and 1223 fatalities in subjects who took the vaccine. Granted, reports to VAERS do not have the vaccine as their sole cause; many people who died after vaccine administration died of causes other than the vaccine. However, this is counterbalanced by underreporting; the VAERS system is thought to have captured less than a tenth of all adverse events.
In short, based on this data, it is entirely possible that the Pfizer vaccine has already killed thousands of people. Who knows how many more will suffer and die from long-term effects from these vaccines? This vaccine injures the myocardium and pericardium, and SARS-CoV-2 Spike inhibits DNA repair and promotes amyloid plaque formation. Are we looking at elevated rates of heart disease, cancer, and neurodegeneration within the next ten years, in the billions of people who have already been inoculated? Nobody knows for certain.
As this is an ongoing event, more data is emerging and more documents are being released with each passing day.
Antibody-Dependent Enhancement
There is another mechanism by which vaccines may fail, and it is known as ADE, or antibody-dependent enhancement of disease. This is something that has been known to occur in Dengue fever. If someone is infected with one strain of Dengue, and then recovers, and is infected with a different strain, their own antibodies can make them very sick. This happens because of something called Original Antigenic Sin. The antibodies from one serotype of Dengue are incompatible with other serotypes. They incompletely neutralize the pathogen, allowing the virus to infect macrophages through the Fc pathway.
Vaccine failure due to ADE has a precedent in recent history. It is thought to be the reason for the failure of Sanofi’s Dengvaxia.
CIDRAP - Sanofi restricts dengue vaccine but downplays antibody enhancement
Pioneering dengue researchers Scott Halstead, MD, and Philip K. Russell, MD, PhD, predicted this move nearly 2 years ago. They had seen results from Dengvaxia's trials showing evidence of antibody-dependent enhancement (ADE), a dengue phenomenon that makes repeat infections more severe and can cause severe illness after vaccination in those who haven't been previously exposed to the virus.
Around the time of the COVID-19 outbreak, Shi Zhengli, the “Bat Lady” at the Wuhan Institute of Virology, published this paper, stating that MERS-CoV was capable of inducing ADE:
Journal of Virology - Molecular Mechanism for Antibody-Dependent Enhancement of Coronavirus Entry
To investigate ADE of coronavirus entry, we first characterized the interactions between Mersmab1 (which is a MERS-CoV RBD-specific MAb) and MERS-CoV spike using biochemical methods. First, enzyme-linked immunosorbent assay (ELISA) was performed between Mermab1 and MERS-CoV RBD and between Mersmab1 and MERS-CoV spike ectodomain (S-e) (Fig. 2A). To this end, Mersmab1 (which was in excess) was used to coat the ELISA plate, and gradient amounts of recombinant RBD or S-e were added for detection of potential binding to Mersmab1. The results showed that both the RBD and S-e bound to Mersmab1. S-e bound to Mersmab1 more tightly than the RBD did, likely due to the multivalent effects associated with the trimeric state of S-e. Second, we prepared Fab from Mersmab1 using papain digestion and examined the binding between Fab and S-e using ELISA. Recombinant S-e (which was in excess) was used to coat the ELISA plate, and gradient amounts of Fab or Mersmab1 were added for detection of potential binding to S-e. The results showed that both Fab and Mersmab1 bound to S-e (Fig. 2B). Mersmab1 bound to S-e more tightly than Fab did, also likely due to the multivalent effects associated with the dimeric state of Mersmab1. Third, a flow cytometry assay was carried out to detect the binding between S-e and the DPP4 receptor and among S-e, Mersmab1, and CD32A (which is an Fc receptor). To this end, DPP4 or CD32A was expressed on the surface of human HEK293T cells (human kidney cells), and recombinant S-e was added for detection of potential binding to one of the two receptors in the absence or presence of Mersmab1. The results showed that without Mersmab1, S-e bound to DPP4 only; in the presence of Mersmab1, S-e bound to CD32A (Fig. 2C). As a negative control, a SARS-CoV RBD-specific MAb (49) did not mediate the binding of S-e to CD32A. The cell surface expressions of both DPP4 and CD32A were measured and used for calibrating the flow cytometry result (Fig. 2D), demonstrating that the direct binding of S-e to DPP4 is stronger than the indirect binding of S-e to CD32A through Mersmab1. Overall, these biochemical results reveal that Mersmab1 not only directly binds to the RBD region of MERS-CoV S-e but also mediates the indirect binding interactions between MERS-CoV S-e and the Fc receptor.
ADE has not been known to occur with SARS-CoV-2, however. On the other hand, it may be that it has yet to appear. With further mutations, antibodies to existing strains may become non-neutralizing.
mRNA-based vaccines provide effective protection against most common SARS-CoV-2 variants. However, identifying likely breakthrough variants is critical for future vaccine development. Here, we found that the Delta variant completely escaped from anti-N-terminal domain (NTD) neutralizing antibodies, while increasing responsiveness to anti-NTD infectivity-enhancing antibodies. Although Pfizer-BioNTech BNT162b2-immune sera neutralized the Delta variant, when four common mutations were introduced into the receptor binding domain (RBD) of the Delta variant (Delta 4+), some BNT162b2-immune sera lost neutralizing activity and enhanced the infectivity. Unique mutations in the Delta NTD were involved in the enhanced infectivity by the BNT162b2-immune sera. Sera of mice immunized by Delta spike, but not wild-type spike, consistently neutralized the Delta 4+ variant without enhancing infectivity. Given the fact that a Delta variant with three similar RBD mutations has already emerged according to the GISAID database, it is necessary to develop vaccines that protect against such complete breakthrough variants.
Although vaccine manufacturers believe they have been diligent enough to avoid inducing ADE with their COVID-19 vaccines, it is important to keep in mind that the Spike in existing vaccines is based on the Wuhan strain of the virus. It is basically like taking a two-year-old flu shot, at best. At worst, it could be priming the body for the emergence of future ADE, along with all the other adverse effects it causes.
Vaccine Efficacy is Rapidly Declining
Although the vaccines were supposedly effective against the Delta variant, their effectiveness at preventing infection with the Omicron variant is practically nil.
We included 16,087 Omicron-positive cases, 4,261 Delta-positive cases, and 114,087 test-negative controls. VE against symptomatic Delta infection declined from 89% (95%CI, 86-92%) 7-59 days after a second dose to 80% (95%CI, 74-84%) after ≥240 days, but increased to 97% (95%CI, 96-98%) ≥7 days after a third dose. VE against symptomatic Omicron infection was only 36% (95%CI, 24-45%) 7-59 days after a second dose and provided no protection after ≥180 days, but increased to 61% (95%CI, 56-65%) ≥7 days after a third dose. VE against severe outcomes was very high following a third dose for both Delta and Omicron (99% [95%CI, 98-99%] and 95% [95%CI, 87-98%], respectively).
It is ridiculous for governments to continue to compel people to take these vaccines.
It’s even more ridiculous when one considers that the vaccines were never really that effective in the first place.
The stated figures of 95% efficacy for the Pfizer vaccine and 94% for the Moderna are based on Relative Risk Reduction, not Absolute Risk Reduction. Relative Risk Reduction is a figure that assumes that everyone is infected at the same time. Absolute Risk Reduction includes those who were not infected. When the ARR of COVID-19 vaccines is calculated, the result is far, far less impressive-sounding.
The Lancet - COVID-19 vaccine efficacy and effectiveness—the elephant (not) in the room
Vaccine efficacy is generally reported as a relative risk reduction (RRR). It uses the relative risk (RR)—ie, the ratio of attack rates with and without a vaccine—which is expressed as 1–RR. Ranking by reported efficacy gives relative risk reductions of 95% for the Pfizer–BioNTech, 94% for the Moderna–NIH, 91% for the Gamaleya, 67% for the J&J, and 67% for the AstraZeneca–Oxford vaccines. However, RRR should be seen against the background risk of being infected and becoming ill with COVID-19, which varies between populations and over time. Although the RRR considers only participants who could benefit from the vaccine, the absolute risk reduction (ARR), which is the difference between attack rates with and without a vaccine, considers the whole population. ARRs tend to be ignored because they give a much less impressive effect size than RRRs: 1·3% for the AstraZeneca–Oxford, 1·2% for the Moderna–NIH, 1·2% for the J&J, 0·93% for the Gamaleya, and 0·84% for the Pfizer–BioNTech vaccines.
In other words, nearly the whole world’s population was inoculated with various COVID-19 vaccines to reduce their actual risk of illness by less than two percent.
But wait, it may be even worse than that. According to Steve Kirsch testifying to the FDA, the vaccines may actually be killing more people than they’re saving:
Dr. Steve Kirsch Bombshell Testimony at FDA Vaccine Hearing - 3681
The UK’s data shows that the vaccinated are still getting sick with COVID-19:
COVID-19 vaccine surveillance report Week 42
So, what purpose does this vaccine actually have?
A Deal With the Devil
These are the terms under which our governments bought the Pfizer vaccine:
Leaked Document Reveals ‘Shocking’ Terms of Pfizer’s International Vaccine Agreements
A leaked document broken down by Twitter user Ehden reveals the shocking terms of Pfizer’s international COVID-19 vaccine agreements.
Countries that purchase Pfizer’s COVID-19 shot must acknowledge that “Pfizer’s efforts to develop and manufacture the product” are “subject to significant risks and uncertainties.”
In the event that a drug or other treatment comes out that can prevent, treat or cure COVID-19, the agreement stands, and the country must follow through with their vaccine order.
While COVID-19 vaccines are “free” to receive in the U.S., they’re being paid for by taxpayer dollars at a rate of $19.50 per dose — Albania, the leaked contract revealed, paid $12 per dose.
The purchaser of Pfizer’s COVID-19 vaccine must also acknowledge two facts that have largely been brushed under the rug: both their efficacy and risks are unknown.
Purchasers must also “indemnify, defend and hold harmless Pfizer … from and against any and all suits, claims, actions, demands, losses, damages, liabilities, settlements, penalties, fines, costs and expenses … arising out of, relating to, or resulting from the Vaccine.”
PFIZERLEAK: EXPOSING THE PFIZER MANUFACTURING AND SUPPLY AGREEMENT.
Pfizer has been extremely aggressive in trying to protect the details of their international COVID19 vaccine agreements.
Luckily, I've managed to get one.
The other vaccine manufacturers made similar contracts with our governments.
The clinical trials for these vaccines were also highly rushed and fraught with fraud and misconduct.
Were the COVID-19 vaccines rushed? Here's how the vaccines were developed so fast

5. Overlapping phases
Regulators at the Food and Drug Administration and those involved in making these vaccines already had seen scientific results on the mRNA vaccine platform. So researchers could focus their questions on animal models and early human trials so that they were completed more quickly. In some instances, there was an overlap of certain study phases.
The Pfizer Vaccine Trial Was Not Double Blind
What Ed and others haven’t seemed to realize yet is that it wasn’t just a problem at an isolated site: the entire trial wasn’t even designed to be double blind in the first place. It was billed as an “observer blinded” study, meaning ostensibly that the researchers don’t know who is getting the treatment but the recipients do or might. In this case, Pfizer said that the appearance of the liquid in the vaccine vials was different than the placebo vials, so the nurses delivering the injection would know the difference, and the trial subjects might get tipped off or figure it out on their own. The nurse wasn’t supposed to tell any of the other researchers who was who. And if none of the researchers know, then they can call it an “observer blinded” study.
Company That Managed Pfizer Vaccine Trial Sites ‘Falsified Data’: Whistleblower
Brook Jackson, a regional director who was employed by Ventavia Research Group, told British Medical Journal that the company “falsified data, unblinded patients, employed inadequately trained vaccinators, and was slow to follow up on adverse events” reported in the pivotal phase III trial of the Pfizer-BioNTech vaccine.
Despite Jackson repeatedly notifying Ventavia of the problems, the company reportedly took no corrective action. This prompted Jackson to send a complaint by email to the US Food and Drug Administration (FDA) – the country’s top regulatory body – in September 2020. But less than 24 hours later, Ventavia fired her in an apparent reprisal.
In a further surprise, the FDA did not follow up on Jackson’s complaint either. Though the regulator claims it doesn’t have the manpower to actively monitor and investigate all clinical trials, one person told the BMJ she was surprised that the agency didn’t inspect Ventavia even after an employee had filed a complaint.
It would be appalling for any drug to be tested and marketed under these conditions, but for pharmaceutical giants to push poison like this, with the widespread approval of our politicians, is just unthinkable.
Undisclosed Ingredients
Numerous groups, including Ricardo Delgado and La Quinta Columna, Andreas Noack (who sadly was rumored to have lost his life over all of this), and UNIT, have come forward stating that the COVID-19 vaccines contain undisclosed graphene nanoparticles. They have examined the vaccine vials under Raman spectroscopy and scanning electron microscopes, and have come to the determination that they have flakes of graphene of various shapes and sizes inside them.
German Chemist, Dr. Andreas Noack, Found DEAD After Exposing Graphene Hydroxide in Covid ‘Vaccines’
UNIT - Case Briefing Document and Lab Report
That last document is also available over IPFS, in uncensorable form:
ipfs://QmaW7qKcE9Q7iATx3caLWQ8fhRfmdP2JM6rrYn8nLkbK69
Graphene promotes oxidative stress in the body, and may affect the function of neurons. Graphene injected into the brains of mice was known to reduce their threat perception (i.e. it had an anxiolytic effect):
Soothing the symptoms of anxiety with graphene oxide
Graphene oxide interrupts anxiety-related neuron signals without affecting the neurons, or the surrounding cells. In simple terms, it only ‘turns down’ the communications between specific neurons. In a disease where these communications are over-expressed, like PTSD and anxiety, targeting the synapses with graphene oxide is enough to halt the development of this pathological behaviour. This is a type of precision medicine.
Graphene may also form the basis of “neural lace” technology, and be used for monitoring of neural activity.
ACS - Effect of Graphene on Nonneuronal and Neuronal Cell Viability and Stress
The cellular biocompatibility of graphene highlights the potential of graphene to be used in biomedical applications by providing long-term and stable nonneural and neural interfaces, especially in the field of bioelectronics to study neural electrophysiology.
Frontiers - Interfacing Graphene-Based Materials With Neural Cells
The scientific community has witnessed an exponential increase in the applications of graphene and graphene-based materials in a wide range of fields, from engineering to electronics to biotechnologies and biomedical applications. For what concerns neuroscience, the interest raised by these materials is two-fold. On one side, nanosheets made of graphene or graphene derivatives (graphene oxide, or its reduced form) can be used as carriers for drug delivery. Here, an important aspect is to evaluate their toxicity, which strongly depends on flake composition, chemical functionalization and dimensions. On the other side, graphene can be exploited as a substrate for tissue engineering. In this case, conductivity is probably the most relevant amongst the various properties of the different graphene materials, as it may allow to instruct and interrogate neural networks, as well as to drive neural growth and differentiation, which holds a great potential in regenerative medicine. In this review, we try to give a comprehensive view of the accomplishments and new challenges of the field, as well as which in our view are the most exciting directions to take in the immediate future. These include the need to engineer multifunctional nanoparticles (NPs) able to cross the blood-brain-barrier to reach neural cells, and to achieve on-demand delivery of specific drugs. We describe the state-of-the-art in the use of graphene materials to engineer three-dimensional scaffolds to drive neuronal growth and regeneration in vivo, and the possibility of using graphene as a component of hybrid composites/multi-layer organic electronics devices. Last but not least, we address the need of an accurate theoretical modeling of the interface between graphene and biological material, by modeling the interaction of graphene with proteins and cell membranes at the nanoscale, and describing the physical mechanism(s) of charge transfer by which the various graphene materials can influence the excitability and physiology of neural cells.
In other words, if these vaccines contain graphene, as well as a protein capable of compromising the blood-brain barrier and potentially delivering graphene across it, this is highly suspicious for numerous reasons.
But Why Are They Doing This?
Why are our governments compelling billions of innocent people, under threat of job loss and being barred from travel and entertainment venues, to take a highly rushed vaccine, produced with novel and untested technology, which has already been shown to cause a litany of highly injurious adverse effects?
There is no way that they could be ignorant of all of this. At this point, one can almost be certain that our governments’ continued insistence on our inoculation with a literal poison and the ruthless elimination of alt-media contradicting their narrative are acts of willful and deliberate malice.
Why would they do this to us, you say? Well, your guess is as good as mine.
On the other hand, I do know that the perpetrators have already stated their intentions decades ago.

The removal of these murderous criminals and megalomaniacs from positions of power and authority over us will take tremendous willpower. It is a battle that is already being fought, worldwide, by all of us.
We are all fighting for our lives.
-Spartacus
This article is licensed under CC BY-SA 4.0. To view a copy of this license, visit http://creativecommons.org/licenses/by-sa/4.0/
Belated welcome to Substack, Spartacus!
I look forward to giving this article the attention it deserves, but I just had to comment on the Georgia Guidestones command to “Balance personal rights with social duties.”
That is precisely what the UK government is in the process of attempting to do at this very moment. They are currently reviewing proposed revisions to their human rights laws to shift emphasis from individual rights to the “wider society”—a.k.a. the “greater good” beloved by dictators throughout history. I just published the letter I submitted to the UK government protesting these reforms if you’d like more details:
https://margaretannaalice.substack.com/p/letter-to-the-uk-government/
Thank you Spartacus.🛡️🗡️ I love that you are writing fairly regularly on substack. This is a great comprehensive overview of the vaccines. I havr links to many of these sections in case others want them if/as they realize they've become sick from the vaccine. Now I have it all in one space, expertly written. 👍🏽
And I LOVED that video on the immune system! One thing I've learned from covid is our immune system is way more complex than I thought. That video breaks it down and a fun way that's easier to understand. I bought the book.